Page 62 - Studia Universitatis Hereditati, vol. 5(2) (2017)
P. 62
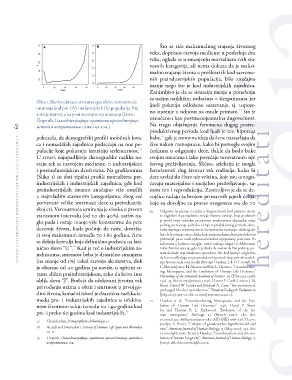
ka 1. Shema obrasca starosno specifične verovatnoćestudia universitatis her editati, letnik 5 (2017), številk a 2 62Što se tiče maksimalnog trajanja životnog
umiranja kod pre- (A) i industrijskih (Б) populacija. Na veka, doprinos razvoja medicine u poslednja dva
x-osi je starost, a na y-osi verovatnoća umiranja (Izvor:hereditati veka, ogleda se u smanjenju mortaliteta svih sta-
Порчић, Палеодемографија: критички преглед теорије, rosnih kategorija, ali nema dokaza da je maksi-
метода и истраживања, слика 42, 104.). malno trajanje života u prošlosti ili kod savreme-
nih preindustrijskih populacija, bilo značajno
pokazala, da demografski profili mobilnih lova- manje nego što je kod industrijskih zajednica.
ca i nomadskih zajednica podsećaju na one po- Zanimljivo je da se situacija menja u poređenju
pulacije koje pokazuju izrazitiju sedentarnost.17 sa našim najbližim rođacima – šimpanzama, jer
U stvari, najupadljivije demografske razlike na- ljudi pokzuju odloženo sazrevanje, tj. uspore-
staju tek sa razvojem medicine, u industrijskim no starenje u odnosu na ostale primate,20 što je
i postindustrijskim društvima. Na grafikonima označeno i kao postmenopauzalna dugovečnost.
(Slika 1) su dati tipični profili mortaliteta pre- Na tragu objašnjenja fenomena dugog postre-
industrijskih i industrijskih zajednica, gde kod produktivnog perioda kod ljudi je tzv. hipoteza
preindustrijskih imamo značajno više umrlih babe,21 gde je osnovna ideja da žene nastavljaju da
u najmlađim starosnim kategorijama, zbog već žive nakon menopauze, kako bi pomogle svojim
pomenute velike smrtnosti dece u preindustrij- ćerkama u odgajanju dece, dakle da budu bake
skoj eri. Verovatnoća umiranja je visoka u prvom svojim unucima i tako povećaju verovatnoću nji-
starosnom intervalu (od 20 do 40%), zatim na- hovog preživljavanja. Slično, selekcija je mogla
glo pada i ostaje manje-više konstantna do pete favorizovati dug životni vek roditelja, kako bi
decenije života, kada počinje da raste, dostižu- dete savladalo čitav niz veština, koje mu omogu-
ći svoj maksimum između 70 i 80 godina, čime ćavaju materijalno i socijalno preživljavanje, sa-
se dobija krivulja koja delimično podseća na lati- mim tim i reprodukciju. Zanimljivo je da se de-
nično slovo “U”.18 Kad je reč o industrijskim za- vojčice rađaju sa brojem primarnih jajnih ćelija,
jednicama, smrtnost beba je drastično smanjena koje su dovoljne za proces ovogeneze sve do 70
(na manje od 1%) usled razvoja akušerstva, dok
je obrazac od 40 godina pa naviše, u opštim cr- 20 Odgovor na pitanje o razlici u dugovečnosti kod primata može
tama sličan preindustrijskom, tako da kriva ima se sagledati iz perspektive teorije životne istorije, koja predstavl-
oblik slova “J”. Budući da očekivani životni vek ja profil vrste određen prosečnim vrednostima dinamike rasta,
pri rođenju uzima u obzir i smrtnost u prvoj go- polnog sazrevanja, početka i kraja reproduktivnog perioda, dina-
dini života, konačni ishod je drastična razlika iz- mike starenja i vremena smrti. Sa stanovišta evolucije, selekcija de-
među pre- i industrijskih zajednica u očekiva- luje da favorizuje one jedinke koje imaju maksimalni reproduktivni
nom životnom veku: između 20 i 40 godina kod potencijal, pa se onda njihova učestalost u populaciji povećava. Po
pre- i preko 60 godina kod industrijskih.19 zakonima o balansu energije, nema razloga ulagati u oblikovanje
takve životne istorije, gde će jedinka da nastavi da živi pošto je pre-
17 Chamberlain, Demography in Archaeology, 11. stala da bude reproduktivno sposobna. Ali, kod ljudi je očigledno
18 Acsádi and Nemeskéri, History of Human Life Span and Mortality, da žene nadživljuju svoj reproduktivni period i da je prirodna selek-
cija favorizovala ovaj model (Kristen Hawkes, J. F. O’Connell, N.
26–8. G. Blurton Jones, H. Alvarez and Eric L. Charnov. “Grandmother-
19 Порчић, Палеодемографија: критички преглед теорије, метода и ing, Menopause, and the Evolution of Human Life Histories.”
Proceedings of the National Academy of Sciences 95 (February 1998):
истраживања, 104. 1336–39, doi:10.1073/pnas.95.3.1336.; Darren P. Croft, Lauren J. N.
Brent, Daniel W. Franks and Michael A. Cant, “The evolution of
prolonged life after reproduction,” Trends in Ecology & Evolution 30
(July, 2015): 407–16, doi: 10.1016/j.tree.2015.04.011.).
21 Hawkes et al., “Grandmothering, Menopause, and the Evo-
lution of Human Life Histories,” 1336.; Daryl P. Shan-
ley and Thomas B. L. Kirkwood, “Evolution of the hu-
man menopause,” BioEssays 23 (March 2001): 282, doi:
10.1002/1521-1878(200103)23:3<282::AID-BIES1038>3.0.CO;2-9.;
Jocelyn. S. Peccei, “Critique of grandmother hypothesis:old and
new,” American Journal of Human Biology 13 (May 2001): 434, doi:
10.1002/ajhb.1076.; Kristen Hawkes, “Grandmothers and the evo-
lution of human longevity,” American Journal of Human Biology 15
(2003): 386, doi:10.1002/ajhb.10156.
umiranja kod pre- (A) i industrijskih (Б) populacija. Na veka, doprinos razvoja medicine u poslednja dva
x-osi je starost, a na y-osi verovatnoća umiranja (Izvor:hereditati veka, ogleda se u smanjenju mortaliteta svih sta-
Порчић, Палеодемографија: критички преглед теорије, rosnih kategorija, ali nema dokaza da je maksi-
метода и истраживања, слика 42, 104.). malno trajanje života u prošlosti ili kod savreme-
nih preindustrijskih populacija, bilo značajno
pokazala, da demografski profili mobilnih lova- manje nego što je kod industrijskih zajednica.
ca i nomadskih zajednica podsećaju na one po- Zanimljivo je da se situacija menja u poređenju
pulacije koje pokazuju izrazitiju sedentarnost.17 sa našim najbližim rođacima – šimpanzama, jer
U stvari, najupadljivije demografske razlike na- ljudi pokzuju odloženo sazrevanje, tj. uspore-
staju tek sa razvojem medicine, u industrijskim no starenje u odnosu na ostale primate,20 što je
i postindustrijskim društvima. Na grafikonima označeno i kao postmenopauzalna dugovečnost.
(Slika 1) su dati tipični profili mortaliteta pre- Na tragu objašnjenja fenomena dugog postre-
industrijskih i industrijskih zajednica, gde kod produktivnog perioda kod ljudi je tzv. hipoteza
preindustrijskih imamo značajno više umrlih babe,21 gde je osnovna ideja da žene nastavljaju da
u najmlađim starosnim kategorijama, zbog već žive nakon menopauze, kako bi pomogle svojim
pomenute velike smrtnosti dece u preindustrij- ćerkama u odgajanju dece, dakle da budu bake
skoj eri. Verovatnoća umiranja je visoka u prvom svojim unucima i tako povećaju verovatnoću nji-
starosnom intervalu (od 20 do 40%), zatim na- hovog preživljavanja. Slično, selekcija je mogla
glo pada i ostaje manje-više konstantna do pete favorizovati dug životni vek roditelja, kako bi
decenije života, kada počinje da raste, dostižu- dete savladalo čitav niz veština, koje mu omogu-
ći svoj maksimum između 70 i 80 godina, čime ćavaju materijalno i socijalno preživljavanje, sa-
se dobija krivulja koja delimično podseća na lati- mim tim i reprodukciju. Zanimljivo je da se de-
nično slovo “U”.18 Kad je reč o industrijskim za- vojčice rađaju sa brojem primarnih jajnih ćelija,
jednicama, smrtnost beba je drastično smanjena koje su dovoljne za proces ovogeneze sve do 70
(na manje od 1%) usled razvoja akušerstva, dok
je obrazac od 40 godina pa naviše, u opštim cr- 20 Odgovor na pitanje o razlici u dugovečnosti kod primata može
tama sličan preindustrijskom, tako da kriva ima se sagledati iz perspektive teorije životne istorije, koja predstavl-
oblik slova “J”. Budući da očekivani životni vek ja profil vrste određen prosečnim vrednostima dinamike rasta,
pri rođenju uzima u obzir i smrtnost u prvoj go- polnog sazrevanja, početka i kraja reproduktivnog perioda, dina-
dini života, konačni ishod je drastična razlika iz- mike starenja i vremena smrti. Sa stanovišta evolucije, selekcija de-
među pre- i industrijskih zajednica u očekiva- luje da favorizuje one jedinke koje imaju maksimalni reproduktivni
nom životnom veku: između 20 i 40 godina kod potencijal, pa se onda njihova učestalost u populaciji povećava. Po
pre- i preko 60 godina kod industrijskih.19 zakonima o balansu energije, nema razloga ulagati u oblikovanje
takve životne istorije, gde će jedinka da nastavi da živi pošto je pre-
17 Chamberlain, Demography in Archaeology, 11. stala da bude reproduktivno sposobna. Ali, kod ljudi je očigledno
18 Acsádi and Nemeskéri, History of Human Life Span and Mortality, da žene nadživljuju svoj reproduktivni period i da je prirodna selek-
cija favorizovala ovaj model (Kristen Hawkes, J. F. O’Connell, N.
26–8. G. Blurton Jones, H. Alvarez and Eric L. Charnov. “Grandmother-
19 Порчић, Палеодемографија: критички преглед теорије, метода и ing, Menopause, and the Evolution of Human Life Histories.”
Proceedings of the National Academy of Sciences 95 (February 1998):
истраживања, 104. 1336–39, doi:10.1073/pnas.95.3.1336.; Darren P. Croft, Lauren J. N.
Brent, Daniel W. Franks and Michael A. Cant, “The evolution of
prolonged life after reproduction,” Trends in Ecology & Evolution 30
(July, 2015): 407–16, doi: 10.1016/j.tree.2015.04.011.).
21 Hawkes et al., “Grandmothering, Menopause, and the Evo-
lution of Human Life Histories,” 1336.; Daryl P. Shan-
ley and Thomas B. L. Kirkwood, “Evolution of the hu-
man menopause,” BioEssays 23 (March 2001): 282, doi:
10.1002/1521-1878(200103)23:3<282::AID-BIES1038>3.0.CO;2-9.;
Jocelyn. S. Peccei, “Critique of grandmother hypothesis:old and
new,” American Journal of Human Biology 13 (May 2001): 434, doi:
10.1002/ajhb.1076.; Kristen Hawkes, “Grandmothers and the evo-
lution of human longevity,” American Journal of Human Biology 15
(2003): 386, doi:10.1002/ajhb.10156.